|
La
corsa nell'uomo: una visione d'insieme bioenergetica
e biomeccanica
di
Gian Nicola Bisciotti (1) (2), Sandra Greco (1),
Claudio Gaudino(1), Jean Marcel Sagnol(3).
1)
Istituto Superiore di Educazione Fisica di Torino
(I).
2) Laboratorio
di Scienza dello Sport UFR-STAPS, Université
de Franche Compté, Besançon (F).
3) Laboratoire
Entraînement et Performance UFR-STAPS, Université
Claude Bernard, Lyon (F).
Abstract:
La corsa, come del resto ogni attività
motoria umana, necessita, per una sua giusta comprensione,
di una visione d’insieme allo stesso tempo
bioenergetica e biomeccanica, questo approccio
multidisciplinare può sottolineare degli
aspetti fondamentali che, al contrario, possono
essere insufficientemente evidenziati quando si
adotti una visione monolaterale del problema.
Questo
tipo di analisi con le sue conseguenze interpretative
può rivelarsi utile, non soltanto nell’ambito
della ricerca ma anche, in funzione della razionalizzazione
della tecnica, nel campo della metodologia di
allenamento.
Introduzione
La
corsa, nonostante sia per l’uomo un’attività
istintiva e naturale, presenta un’obbiettiva
complessità meccanica, in una sua prima
analisi, sia pur ridotta e semplificata, possiamo
notare come l’energia cinetica (Ecin = _
M · V2) e l’energia potenziale (Epot
= m·g ·h) risultino in fase tra loro
e come altresì durante la fase di volo
nessun tipo di forza esterna, essendo la resistenza
dell’aria trascurabile, possa apportare delle
modifiche alla bioenergetica del moto stesso.
Inoltre
nelle prima parte della fase di volo Epot aumenta
a causa della diminuzione dell’ Ecin e diminuisce
nella seconda parte quando a sua volta si tramuta
in Ecin.
Infine
nel momento in cui il piede prende contatto al
suolo, si ha una diminuzione dell’energia
totale (Etot) del sistema poiché si verifica
un abbassamento del centro di gravità simultaneamente
alla decelerazione.
Da
un punto di vista puramente meccanico quindi la
corsa è assimilabile alla successione di
rimbalzi di un corpo elastico in cui si possono
distinguere una fase aerea effettiva ed un tempo
di contatto effettivo.
La
meccanica della corsa può quindi essere
esemplificata attraverso l’immagine di un
pallone che rimbalza al suolo, come è possibile
vedere nella Fig. 1.
Nel
momento in cui il pallone tocca terra l’Ecin
e l’Epot scendono simultaneamente a zero,
il pallone risulta infatti fermo per un tempo
minimo, in questa fase una parte del lavoro meccanico
negativo , imputabile essenzialmente alla deformazione
elastica della palla, viene accumulato sotto forma
di energia elastica che verrà susseguentemente
restituita nel corso del rimbalzo successivo dando
luogo in tal modo ad un simultaneo aumento dell’
Ecin e dell’Epot.
Per
questo motivo nella corsa, esattamente come nel
rimbalzo di un pallone al suolo, non vi può
essere alcuna trasformazione di Ecin in Epot e
viceversa, tuttavia la fase di accumulo e di conseguente
restituzione di energia elastica consente un importante
risparmio energetico.
Infine
è importante notare come a basse velocità
di corsa il rimbalzo del corpo sia sostanzialmente
simmetrico, ossia la durata del tempo di contatto
effettivo è pari alla durata della fase
aerea effettiva, mentre ad alte velocità
di corsa tale rimbalzo diviene asimmetrico a causa
dell’aumento temporale della fase aerea (Cavagna
e coll. 1988 )
La
frequenza e l’ampiezza del passo
Il
controllo della frequenza e dell’ampiezza
del passo sono parametri fondamentali nella valutazione
dell’andamento generale della corsa, e costituiscono
un aspetto assai importante al fine di ottenere
un tipo di corsa il più economico possibile
che permetta di realizzare la più alta
velocità media in rapporto alla distanza
da percorrere. Il concetto di frequenza può
essere definito come il numero di movimenti completi
compiuti da un punto P nell’unità
di tempo (f=1\t), che nel caso specifico della
corsa diviene f=1/( Tc+Tv ),
dove Tc è il tempo di contatto
del piede al suolo e Tv il tempo
di volo; tale valore risulta dipendente sia dal
numero dei passi che dal tempo considerato.
La
velocità di corsa (V) è il risultato
del prodotto della lunghezza (SL) e della frequenza
del passo (SF), V = SL·SF.
Quando
la velocità di corsa aumenta, il soggetto
tende ad aumentare la lunghezza della falcata
e la sua frequenza di passo, ma mentre
SL, pur aumentando in funzione della velocità
di corsa, raggiunge un valore stabile ad alte
velocità di percorrenza, SF cresce in modo
relativamente maggiore a velocità di corsa
considerevoli (Williams 1985).
I
fattori che determinano questo tipo di adattamento
sembrano sostanzialmente incentrati sulla ricerca
di una massima economia di corsa basata su di
un ottimale frequenza del passo (Dalleau e coll.
1998a,b), questa teoria era già stata avanzata
sia da Högberg (1952) è più
recentemente da Cavanagh e Williams (1982) che
mostrarono come la frequenza spontanea fosse simile
alla frequenza che presenta il valore minimo sulla
curva del costo metabolico in funzione della frequenza
del passo stesso.
Altri
Autori hanno suggerito come SF fosse scelta in
modo tale da minimizzare, per una data velocità
di corsa, la potenza meccanica totale, precisando
ulteriormente come in effetti SF abbia effetti
opposti sulla potenza meccanica esterna e su quella
interna (1), la prima infatti decresce con l’aumentare
della frequenza, mentre la seconda al contrario
aumenterebbe di valore (Kaneko e coll.1987).
Recentemente
Cavagna e coll. (1991) approfondendo la problematica
di studio legata agli effetti del cambiamento
della frequenza sulla produzione di potenza meccanica,
hanno dimostrato come SF spontanea tenda a minimizzare
la potenza meccanica prodotta durante un passo
effettuato a bassa velocità di corsa, mentre
ad elevate velocità di percorrenza tenda
a minimizzare unicamente la potenza meccanica
della fase propulsiva.
Inoltre
è da sottolineare come la stiffness degli
arti inferiori, ossia la loro rigidità
muscolo-tendinea, aumenti linearmente in funzione
della frequenza dei passi (Farley e Gonzales 1996)
provocando in tal modo un aumento della trasmissione
degli shocks generati dall’impatto del piede
al suolo (Mc Mahon 1984, Mc Mahon e coll. 1987,
Lafortune e coll. 1996).
In
seguito affronteremo la tematica inerente a come,
durante la corsa, il soggetto cerchi di adattare
la frequenza del passo sia in funzione di un’ottimale
economia di corsa che di una minimizzazione degli
shocks trasmessi all’apparato osteo-muscolo-legamentoso.
Anche
la lunghezza del passo riveste un ruolo determinate
nel dispendio energetico e nel rendimento della
corsa, infatti il VO2 per un data velocità
di corsa aumenta in maniera curvilinea rispetto
all’aumento od alla diminuzione della lunghezza
del passo naturalmente scelta dall’atleta
(Höeberg 1952, Cavanagh e Williams 1982),
recenti studi hanno inoltre dimostrato come attraverso
la correzione effettuata tramite retrocontrollo
audiovisuale di una lunghezza di passo non ottimale
(ossia non corrispondente ad una minimo consumo
di O2 ) si sia potuta ottenere una diminuzione
della richiesta energetica della corsa stessa
(Morgan e coll. 1994).
Rendimento
e costo energetico della corsa
Il
rendimento muscolare in situazioni di attivazione
naturale risulta essere di circa il 25% (Cavagna
e coll. 1964), tuttavia nella corsa il rendimento
raggiunge percentuali di circa 40-50%, mentre
in altre attività muscolari come il ciclismo
questa percentuale raggiunge solamente circa il
20% (Gaesser e Brooks,1975)
Questo
aumentato rendimento muscolare nel caso della
corsa sembra essere attribuibile alla restituzione
di energia elastica che avviene durante la biomeccanica
della corsa stessa.
Nella
fase eccentrica infatti viene accumulata energia
elastica nella componente elastica seriale (essenzialmente
il tendine e la parte S2 della miosina) della
muscolatura che viene sottoposta ad allungamento,
energia che viene poi restituita sotto forma di
lavoro meccanico potenziando la successiva fase
concentrica.
Il
recupero dell’energia elastica immagazzinata
nella fase eccentrica permetterebbe quindi una
diminuzione della spesa energetica (Cavagna e
coll. 1964, Cavagna e coll. 1968, Shorten 1987)
inoltre il ruolo svolto dal recupero di energia
elastica sarebbe ancor più rilevante a
velocità di corsa elevate (Bosco e Rusko
1983).
Il
fenomeno di stoccaggio e restituzione di energia
elastica è stato confermato sperimentalmente
da misurazioni effettuate con l’ausilio della
piattaforma di forza, dove è riscontrabile
come la forza di reazione al suolo sia una funzione
lineare dello spostamento del corpo (Cavagna e
coll.1988) ed è proprio questa linearità
che suggerisce come il comportamento dell’arto
di appoggio sia simile a quello di una molla,
concetto che descriveremo successivamente in modo
più specifico nello Spring Mass Model (Alexander
1988) .
Il
costo energetico della corsa (Cr),
si ottiene dividendo il VO2 netto,
corrispondente ad una velocità sottomassimale
determinata, per la velocità stessa (Margaria,
1938;Margaria e coll.,1963) :
Cr
= (VO2 - VO2 riposo)
· v-1
Dove
V O2 è espresso in ml ·
kg ·s-1 e v è la velocità
di corsa espressa in m · s-1,
in tal modo Cr risulta essere espresso
in ml di O2 oppure in Joules per kg
di massa corporea e per metro percorso.
Tale
valore rimane comunque legato a delle costanti
individuali molto specifiche, tuttavia il Cr
si rivela indipendente rispetto alla velocità
di corsa sino a valori di circa 6 m · s-1
(Åstrand, 1952; Krahenbuhl e coll,1979;
Conley e Krahenbuhl, 1980; Davies, 1980; Brandsford
e Howley, 1977; Di Prampero e coll., 1986 e 1993)
e risulta essere di circa 0.9 kcal
· kg-1 · km-1
per tale motivo è possibile paragonare
il Cr interindividuale a diverse velocità
di percorrenza .
Tuttavia
questo tipo di calcolo del dispendio energetico
risulta affidabile soltanto nel caso di velocità
di percorrenza costante, in altre parole il dispendio
energetico della corsa risulta essere di questo
tipo solamente in caso di assenza di fasi rilevanti
di accelerazione e decelerazione.
Infatti,
ipotizzando che il costo dell’accelerazione
sia uguale a _ M V2 max
( dove M è la massa dell’atleta e
V max la velocità massima raggiunta
) ( Di Prampero 1985) ed il rendimento muscolare
pari a circa il 25% ( Cavagna 1988 ) è
possibile calcolare indirettamente il costo energetico
supplementare relativo alle fasi di accelerazione
eventualmente presenti nella corsa stessa.
L’esempio
seguente potrà chiarire maggiormente il
concetto sovraesposto:
Supponendo
che un atleta corra reiteratamente un tratto di
20 m.t. a velocità sub-massimale (compresa
tra 2 e 4 m.t · s-1 ) in corsa-navette, ovverosia
invertendo la direzione di corsa ogni 20 m.t.
di percorrenza attraverso un cambio di direzione
seguito da una fase di accelerazione stimata di
una lunghezza di circa 2.5 m.t., il rapporto tra
la sua velocità massimale (Vmax) e la sua
velocità media (Vmed) può essere
calcolato come segue:
Vmed
= (2.5 Vmax)/2 + 15 Vmax + (2.5 Vmax)/2
20
da
cui : Vmax = 8/7 Vmed
Con
lo stesso tipo di calcolo è possibile stimare
il rapporto tra Vmax e Vmed se la stessa quantità
totale di corsa viene effettuata a parità
di tempo ma invertendo la direzione di corsa ogni
10 m.t. (navette 10 m.t.) od ogni 5 m.t. (navette
5 m.t.) ossia rispettivamente raddoppiando e quadruplicando
i cambi di direzione e conseguentemente le fasi
di accelerazione :
Nel
caso di due tratti da 10 m.t. avremo:
Vmed
= (2.5 Vmax)/2 + 5 Vmax + (2.5 Vmax)/2
10
da
cui: Vmax = 4/3 Vmed
Nel
caso invece di quattro tratti da 5 m.t. il calcolo
sarà il seguente:
Vmed
= (2.5 Vmax)/2 + (2.5 Vmax)/2
5
da
cui: Vmax = 2 Vmed
Nel
caso di una corsa-navette prolungata con cambio
di direzione ogni 20 m.t. l’energia cinetica
(Ecin) richiesta per le fasi di accelerazione
sarà dunque pari a :
Ecin
= _ M V2 = _ M (2 Vmed) 2
Da
cui:
Ecin
= _ M (8\7 Vmed ) 2 = 64/98 M V2 med » 0.65
M V2 med
Nel
caso di una corsa-navette prolungata con cambio
di direzione ogni 10 m.t. l’energia cinetica
(Ecin) richiesta per le fasi di accelerazione
sarà pari a :
Ecin
= _ M V2 = _ M (2 Vmed) 2
Da
cui:
Ecin
= _ M (4/3 Vmed ) 2 = 16/18 M V2 med » 0.89
M V2 med
Infine
nel caso di una corsa-navette prolungata con cambio
di direzione ogni 5 m.t. l’energia cinetica
(Ecin) richiesta per le fasi di accelerazione
sarà pari a :
Ecin
= _ M V2 = _ M (2 Vmed) 2
Da
cui :
Ecin
= 4/2 M V2 med = 2 M V2 med
Riepilogando
quindi nella corsa-navetta prolungata con cambio
di direzione ogni 20 m.t., Vmax sarà uguale
a 8/7 Vmed e l’Ecin = 64/98 MV2 med.
Nel
caso di una corsa-navetta prolungata con cambio
di direzione ogni 10 m.t., Vmax sarà uguale
a:
4/3
Vmed ed Ecin = 16/18 MV2 med.
Infine
nel caso di una corsa-navetta prolungata con cambio
di direzione ogni 5 m.t., Vmax sarà uguale
a 2 Vmed ed Ecin = 2 MV2 med.
Assumendo
che il costo metabolico della corsa nell’uomo
sia pari a circa 0.9 Kcal · kg-1 · km,
e trasformando il costo metabolico della presente
in Joule (essendo 1 Kcal pari a 4,18 kJ ), avremo
un costo energetico per metro pari a circa :
4
J · M
nella
quale M è la massa del soggetto espressa
in kg.
Da
cui potremmo calcolare che il costo energetico
sui diversi tipi di frazionamento di corsa-navetta:
corsa
frazionata su tratti di 5 m.t. : J » 20 ·
M
corsa
frazionata su tratti di 10 m.t. : J » 40
· M
corsa
frazionata su tratti di 20 m.t. : J » 80
· M
Ipotizzando
ad esempio una velocità di percorrenza
media pari a 2 m.t. · sec —1 ed un rendimento
muscolare pari al 25%, il costo energetico necessario
alle fasi di accelerazione di una corsa-navetta
frazionata in tratti da 5 m.t. sarà pari
a:
Ecin
= 2 M · (2)2 / 0.25 =32 M
Nel
caso di una corsa-navetta frazionata in tratti
da 10 m.t. il costo energetico inerente alle fasi
di accelerazione sarà pari a:
Ecin
= 16/18 M · (2)2 / 0.25 = 14,22 M
Infine
nel caso di una corsa-navetta frazionata in tratti
da 20 m.t. il costo energetico inerente alle fasi
di accelerazione sarà pari a:
Ecin
= 64/98 M · (2)2 / 0.25 = 10.45 M
Per
cui il costo totale (Etot) di una corsa prolungata
frazionata in tratti-navetta di 5 m.t. percorsi
ad una Vmed di 2 m.t. · s-1 sarà pari
a :
Etot
= 10 M +32 M = 42 M
per
cui ogni tratto totale di 20 m.t. comporterà
un dispendio totale di 168 J ·kg-1
Nel
caso di tratti-navetta di 10 m.t. sarà
pari invece a :
Etot
= 30 M + 14.22 M = 44.22 M
per
cui ogni tratto totale di 20 m.t. comporterà
un dispendio totale di 88,44 J ·kg-1
Infine
nel caso di corsa-navetta frazionata in tratti
di 20 m.t. si avrà un costo pari a :
Etot
= 70 M + 10,45 M = 80,45 J ·kg-1 per ogni
tratto considerato.
Nel
caso dello stesso tipo di percorrenza effettuato
però ad una velocità doppia (4 m.t.
· sec-1) rispetto all’esempio precedente,
Etot sarà pari a:
Etot
5 m.t. = 10 M + 128 M = 138 M
rapportato
ad un tratto di 20 m.t. quindi
Etot
= 552 J ·kg-1
Etot
10 m.t. = 30 M + 57 M = 87 M
Rapportato
ad un tratto di 20 m.t. quindi
Etot
= 174 J ·kg-1
Etot
20 m.t. = 70 M + 42 M = 112 J ·kg-1
Il
dispendio energetico della corsa quindi sarebbe
fortemente correlato al numero delle fasi di accelerazione
ed alla velocità media da mantenersi in
funzione delle accelerazioni stesse nonché
al cambio di frequenza di passo che tali fasi
di accelerazione e decelerazione comportano, come
vedremo più avanti infatti l’adozione
di una frequenza di passo che si allontani dalla
frequenza naturale, comporta un maggior dispendio
energetico.
Infatti
dai calcoli sopraindicati possiamo desumere come
alla Vmed di 2 m.t. sec-1 il costo metabolico
di un tratto di corsa di 20 m.t. salga di circa
1,1 volte quando lo stesso tratto venga percorso
in due tratti navette di 10 m.t. e di circa 2
volte nel caso in cui venga percorso in quattro
tratti navette di 5 m.t.
Questi
costi salgono ulteriormente a 1.5 ed a 4,9 nel
caso di percorrenza alla Vmed di 4 m.t. sec-1
.
Questi
calcoli teorici possono essere ritenuti sufficientemente
coerenti ai dati registrati nel corso di una sperimentazione
nella quale a 14 giocatori di calcio di medio
livello di età pari a 25 +4 anni
veniva richiesto di effettuare una corsa -navetta
sino ad esaurimento su tratti di 20, 10 e 5 m.t.
a velocità progressivamente crescenti (
comprese tra 2.5 e 3.6 m.t. sec-1 ) secondo il
protocollo di test di Léger e coll. (Leger
e coll. 1982a, 1982b,1984,1985).
I
risultati registrati indicano un decremento della
durata e della velocità massima di percorrenza
pari al 41.01+6.42% tra navette 20 m.t.
e navette 10 m.t., del 73.74+5.31% tra
navette 20 m.t. e navette 5 m.t. e del 54.73+11.47%
nel caso di navette 10 m.t. e navette 5 m.t. (Bisciotti,
Belli 1998).
Tali
risultati, anche se richiedono un ulteriore approfondimento
sperimentale attraverso un protocollo che preveda
la misurazione diretta del consumo di O2, rendono
evidente la difficoltà nella quantificazione
del dispendio energetico in alcune attività
come il calcio, il rugby, la pallacanestro od
il tennis, nelle quali la corsa è caratterizzata
dall’alternarsi di fasi di accelerazione
a fasi corsa blanda a velocità pressoché
costante ed a continui cambi nella frequenza del
passo di corsa dettati dall’esigenza di essere
in sintonia con la traiettoria e la velocità
della palla.
Da
quanto esposto appare quindi chiara la necessità
dell’appropriata metodologia di allenamento
che tali attività richiedano proprio in
virtù della loro specificità bioenergetica.
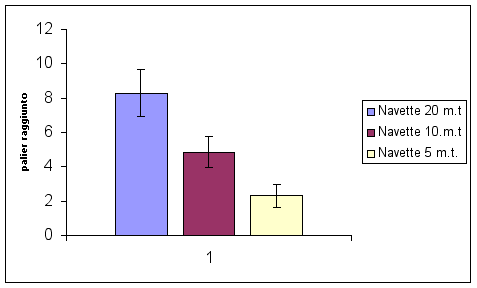
Palier raggiunto in funzione dell’aumento
dei cambi di direzione e delle conseguenti fasi
di accelerazione di 3 differenti modalità
di corsa - navetta (20,10 e 5 m.t.).
Ogni
palier corrisponde ad 1’ di corsa a velocità
progressivamente crescente, per cui maggiore è
il palier raggiunto maggiore risulta essere la
più alta Vmed di percorrenza registrata
durante il test (Bisciotti, Belli 1998).
Il
rapporto tra VO2 e Cr
Il
VO2 max espresso in ml·kg-1
è sempre stato considerato uno dei parametri
maggiormente discriminanti nell’ambito della
performance della corsa prolungata (Saltin e Åstrand
, 1967; Costil e coll., 1973 ; Boileau e coll.,
1982).
Il
Cr rimane tuttavia correlato negativamente
rispetto al peso e all’altezza individuale,
dal momento che questi due fattori sono difficilmente
discriminabili per poter valutare coerentemente
il VO2 max indipendentemente
dalla massa, occorrerebbe dare a quest’ultima
un esponente uguale a 0,75 (Svendenhag e Sjödin,
1994) apparirebbe più corretto quindi legare
il concetto di rendimento di corsa a quello di
VO2 ·ml-1·kg-0.75
, in tal modo il VO2 (lt ·
min-1) sarebbe proporzionale alla massa
potenza di 2/3.
Tuttavia
il VO2 max non è il solo elemento
esplicativo nelle differenze interindividuali
del rendimento della bioenergetica della corsa,
in effetti il fattore maggiormente discriminante
nei confronti della massima velocità di
corsa sostenibile sembrerebbe essere attribuibile
al Cr .
In
effetti un miglioramento del 10% del Cr
comporterebbe un aumento del 12% della Velocità
Aerobica Massimale (Va max, ) che a
sua volta consentirebbe un aumento nella velocità
massimale di corsa pari al 9% nel corso di una
prestazione di 5.000 m.t. (Lacour, 1990).
La
Va max, concetto introdotto da Di Prampero
nel 1986, è ottenibile attraverso l’applicazione
della seguente formula :
Va
max = (VO2 max
- 0.083) · Cr-1
Dove
VO2 è espresso in ml ·
kg-1 · s-1, v in m · s-1 ed il valore
di 0.083 ml · kg -1· s-1 ( 5 ml ·
kg-1 · m-1 ), stabilito da Medbø e
coll (1988), è la media dell’intercettazione
dell’asse y nella relazione lineare VO2
· v-1ottenuta durante la corsa su treadmill
e corrisponde al VO2 a riposo dell’atleta
in piedi sul treadmill stesso.
Dal
momento che le possibilità di aumento del
Cr nella bioenergetica individuale di corsa sono
comprese in un range che va dal 5 al 16% del Cr
totale (Conley e coll., 1981 e Conley e coll.,
1984), è importante ricordare l’importanza
che riveste il Cr nell’economia
totale di corsa.
Lo
Spring Mass Model
Come
già precedentemente ricordato, nella biomeccanica
e nella bioenergetica della corsa l’elasticità
muscolare svolge un ruolo molto importante, per
questo motivo nell’ambito della modellizzazione
della corsa sia nell’uomo che in alcuni tipi
di animali, come ad esempio il canguro od il montone,
si è arrivati alla concettualizzazione
di un modello meccanico costituito da una massa
, che riveste il ruolo della massa corporea totale
in serie ad una componente elastica , sotto forma
di una molla, che modellizza gli arti inferiori
e le loro caratteristiche elastiche.
Nell’interpretazione
concettuale di tale modello un elemento esplicativo
della variazione interindividuale del Cr
nella bioenergetica della corsa umana, può
essere attribuito alla stiffness neuromuscolare.
La
stiffness neuromuscolare è definibile come
una variazione di forza su una variazione di lunghezza
ed è quantificabile matematicamente come
il rapporto di D F/D L nel quale D
L è una funzione lineare della forza D
F (Cavagna,1988).
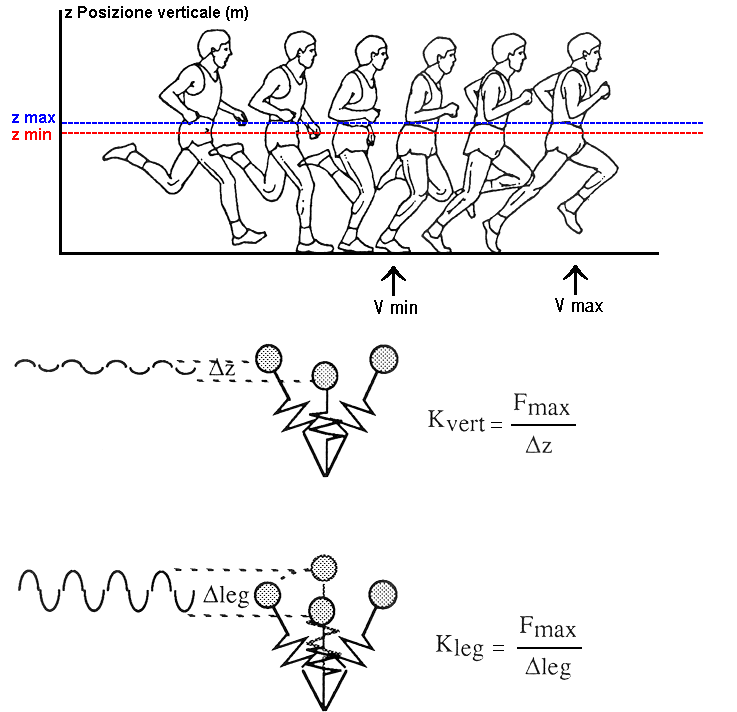
Figura
1: lo Spring Mass Model applicato alla biomeccanica
della corsa (Mc Mahon e Cheng, 1990). In questo
tipo di modello, se si osservano solamente le
oscillazioni sul piano verticale del corridore,
si parla di stiffness verticale (Kvert),
mentre se si tiene conto del reale accorciamento
della gamba si parla di stiffness dell’arto
inferiore (Kleg)
Lo
Spring Mass Model (Alexander e Vernon, 1975) inizialmente
concepito per lo studio del meccanismo di stoccaggio
e restituzione di energia elastica nella locomozione
del canguro, è stato applicato con successo
anche nell’ambito della locomozione umana
(Alexander, 1988; Blickhan, 1989;Mc Mahon, 1990;Mc
Mahon e Cheng, 1990).
Quando
questo tipo di modellizzazione viene applicato
alla corsa nell’uomo, occorre distinguere
tra Kvert e Kleg ( Mc Mahon
e Cheng 1990 ), intendendo per Kvert
le oscillazioni verticali del corridore, e riferendosi
in questo caso alla stiffness verticale, e per
Kleg il reale accorciamento dell’arto
inferiore d’appoggio, intendendo in tal caso
la stiffness degli arti inferiori.
Per
meglio comprendere il concetto di stiffness occorre
fare riferimento alla "rigidità"
della struttura meccanica considerata, che nel
caso specifico è la struttura muscolo-
tendinea.
Ogni
struttura meccanica quando subisce una perturbazione
della sua posizione di equilibrio, vibra ad una
frequenza variabile che dipende dalla sua massa
e dal suo valore di stiffness K.
La
frequenza di risonanza si può calcolare
applicando la seguente equazione che definisce
la frequenza di un moto armonico:
RF
= 1/ 2p Ö
K/M
Dove
K è la rigidità ed M la massa del
corpo.
Il
valore di rigidità K può essere
quantificato attraverso differenti tipi di calcolo,
il metodo più classico può forse
essere considerato quello di Farley e coll. ( 1991)
che, assimilando il soggetto ad un sistema composto
appunto da una massa e da una molla rimbalzante
al suolo , parte dal presupposto concettuale che
la forza esercitata da tale complesso sul suolo
stesso passi per il suo peso durante il periodo
di contatto e di stacco. La durata tra questi
due istanti, misurata su pedana di forza, è
considerata come la metà del periodo di
risonanza del sistema oscillante (T/2) , e la
rigidità del sistema massa-molla rimbalzante
al suolo e quindi calcolata attraverso la formula :
K
= p 2 / (T/ 2) 2 (in N · m-1
· kg-1)
In
tal modo conoscendo la frequenza reale del passo
(SF), si può calcolare la differenza relativa
D F % risultante dal rapporto tra SF e RF
applicando la seguente formula:
D
F % = (SF — RF)· RF-1 %.
Un
primo dato interessante che occorre sottolineare
è come il Cr risulti inversamente
correlato alla rigidità e direttamente
correlato alla differenza tra SF ed RF, ossia
maggiore risulti questa differenza, maggiore appare
essere il costo energetico della corsa stessa
(Dalleau e coll. 1998a,b).
Sembrerebbe
quindi che la frequenza ideale di passo alla quale
il Cr è minimo, sia prossima
alla frequenza naturale di oscillazione del sistema
(Cavagna e Franzetti 1982, Kaneko e coll. 1987)
In
effetti quando un sistema formato da una massa
ed un corpo elastico, come nel caso dello spring
mass model, viene forzato ad una frequenza RF,
la forza e l’energia necessaria
per realizzare una data ampiezza di movimento
sono minime, in altri termini RF è la frequenza
che permette il mantenimento di un movimento oscillante
ad una data ampiezza con un dispendio energetico
minimo, per questo motivo numerosi autori indicano
come RF possa minimizzare il dispendio energetico
anche nel movimento umano (Mc Mahon 1984, Taylor
1985, Dalleau e coll. 1998a,b).
E’
comunque interessante notare che il valore minimo
di Cr non corrisponda al valore massimo
di rendimento, infatti nel momento in cui la frequenza
aumenta si verifica un aumento del Cr
stesso probabilmente dovuto ad un parallelo aumento
del costo meccanico (
Kaneko 1990 ).
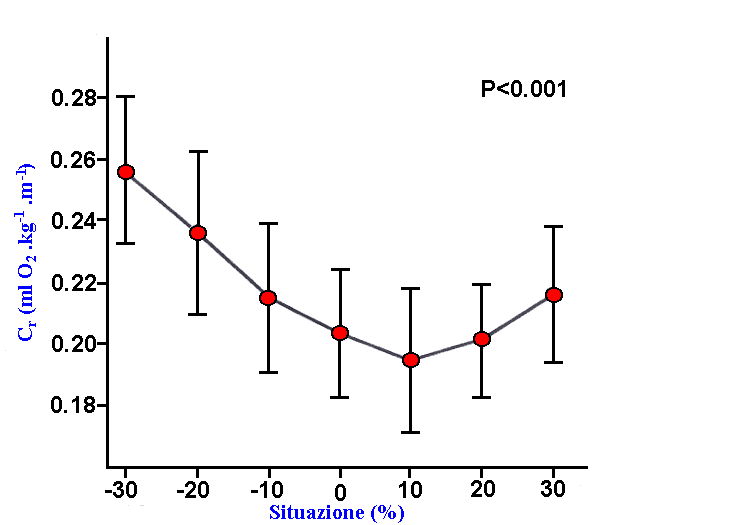
Figura
2: influenza della variazione della frequenza
dei passi sul Cr ( da Dalleau e coll. 1998a, modificato).
Infatti
dal momento che la stiffness degli arti inferiori
risulta essere una funzione lineare della frequenza
dei passi (Farley e Gonzales, 1996 ) ad elevate
frequenze di passo interverrebbe probabilmente
un aumento della stiffness del sistema neuro-muscolare
con un conseguente maggior lavoro meccanico interno
(Dalleau, 1998a), inoltre occorre ricordare come
una stiffness elevata comporterebbe un aumento
dello shock trasmesso al sistema muscolo-scheletrico,
favorendo probabilmente l’insorgenza di lesioni
a carico dell’apparato locomotore (McMahon
1984, McMahon e coll. 1987).
Numerosi
sono infatti gli studi che confermano l’aumento
dei valori di stiffness in rapporto all’aumento
della velocità di percorrenza, Blickhan
(1989) riferisce valori di stiffness che variano
entro i 10 kN · m-1 a 3 m · s -1 e 30
kN · m-1a 9 m·s—1, Farley e Gonzales
(1996) riportano valori di stiffness dell’ordine
di 11 kN· m-1 alla velocità di 2.5
m · s-1, mentre Viale e coll. (1997) riferiscono
valori di 13 kN · m-1 ad una velocità
di percorrenza pari a 4 m · s-1 . infine
Dalleau e coll. (1998a,b) riferiscono come la
stiffness neuro-muscolare raddoppi di valore nel
momento in cui la frequenza passi dal —30%
al +30% della frequenza di passo spontanea, indicando
in tal modo come la stiffness possa dipendere
non solo dall’aumento della velocità
di percorrenza ma anche dalla frequenza.
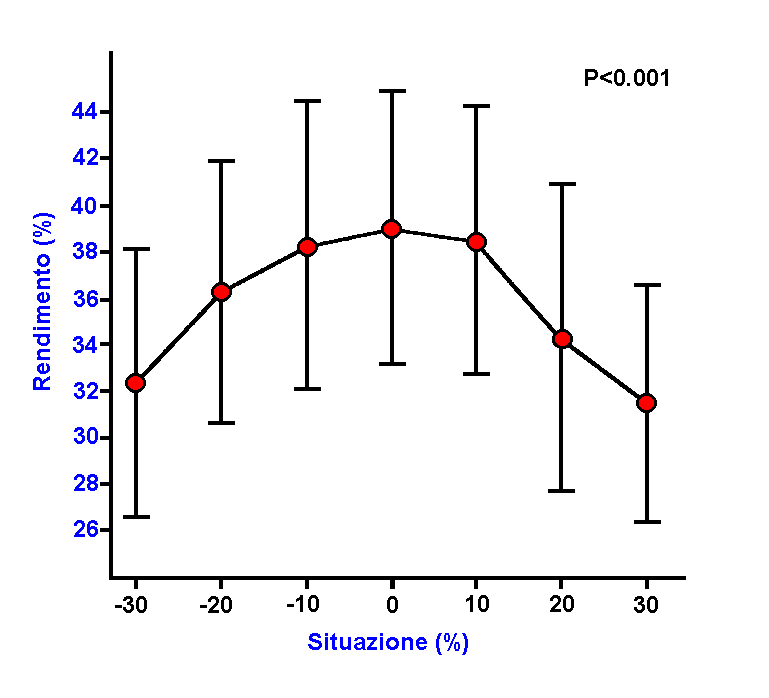
Figura
3: Influenza della variazione della frequenza
dei passi sul rendimento ( da Dalleau e coll.
1998a, modificato.
Inoltre
dal momento che la stiffness degli arti inferiori
è riconducibile sia alla rigidità
muscolo-tendinea dei muscoli estensori della gamba
sia alla struttura elastica del piede (Kerr e
coll.1987, Alexander 1988, Alexander 1997), è
interessante notare come un piede supinato sia
maggiormente rigido di un piede pronato e come
questo comporti un irrigidimento del sistema durante
la corsa (Viale e coll.1998)
E’
importante infine ricordare che la stiffness,
anche se risulta fortemente influenzata sia dalla
rigidità dei ponti acto-miosinici che da
quella dei tendini (Shorten 1987, Alexander 1988),
è modulabile solo attraverso l’attivazione
neuro-muscolare.
Da
quanto sopra esposto quindi sembra che durante
la corsa il massimo rendimento si abbia quando
il soggetto possa modulare la rigidità
del suo apparato muscolo-tendineo in maniera tale
che RF del modello sia il più simile possibile
ad SF reale.
Tuttavia
SF tende ad identificarsi con RF soprattutto a
basse velocità di corsa (Cavagna e coll.
1988) mentre per velocità più elevate,
alle quali interverrebbe un aumento della stiffness
con conseguente maggior shock a carico dell’apparato
locomotore, il soggetto tende ad adottare una
frequenza del 10% inferiore, in tal modo sembra
che l’atleta preferisca una frequenza di
corsa inferiore con un conseguente maggiore Cr,
visto che quest’ultimo presenterebbe il suo
valore minimo ad un valore di frequenza superiore
del 10% rispetto alla frequenza spontanea, probabilmente
al fine di ottimizzare il compromesso tra la necessità
di preservare le proprie strutture anatomiche
e la massimalizzazione della performance legata
al minimo dispendio energetico (Dalleau e coll.
1988a,b).
Alla
luce di queste considerazioni possiamo avanzare
l’ipotesi che la regolazione della stiffness
dell’apparato muscolo-tendineo durante la
corsa, attuabile attraverso una regolazione neuro-muscolare,
possa essere interpretabile anche come un meccanismo
di protezione contro i microtraumatismi ripetuti
dovuti allo shock dell’impatto del piede
al suolo.
Un
ulteriore dato interessante che può emergere
da un’analisi approfondita di questo tipo
di modellizazione muscolare è che calcolando
la rigidità, la frequenza di risonanza
ed il lavoro meccanico di ogni arto separatamente,
possiamo constatare come un arto rispetto al controlaterale
realizzi una maggior quantità di lavoro
meccanico e possa essere per questa ragione definito
"arto propulsivo" dal momento che risulta
essere quello che svolge l’azione propulsiva
orizzontale e verticale maggiore, mentre l’altro
è definibile come arto di appoggio, vista
la sua minore azione di flessione-estensione durante
la fase di supporto (Cavanagh 1990), questa asimmetria
nella produzione di lavoro positivo è quantificabile
in circa il 20% (Dalleau e coll. 1998a,b).
Conclusioni
L’ottimizzazione
dei vari parametri che costituiscono quella che
per l’uomo risulta essere una delle attività
più istintive e naturali quali la corsa
comporta, nel momento in cui la performance venga
estremizzata come nella realtà sportiva
di alto livello, una profonda conoscenza di tutti
i suoi aspetti biomeccanici ed energetici.
Per
questo motivo non solo sarebbe auspicabile una
profonda conoscenza teorica di questi ultimi anche
da parte del tecnico sportivo ma ancor di più
risulterebbe fondamentale una stretta collaborazione
tra il ricercatore e l’ "uomo di campo"
allo scopo di individuare su solide basi scientifiche
le migliori metodologie di allenamento atte al
miglioramento della prestazione onde evitare l’applicazione
di una linea metodologica di lavoro basata su
quel tanto sino ad oggi utilizzato metodo di apprendimento"
per prove ed errori" che nasce da un sostanziale
empirismo valutativo e che non ha più ragione
di esistere nell’ambito della prestazione
sportiva di alto profilo qualitativo.
Nota:
(1) Per potenza meccanica esterna si intende la
potenza relativa agli spostamenti del centro di
gravità del corpo rispetto al suolo e per
potenza meccanica interna quella relativa allo
spostamento degli arti rispetto al centro di gravità.
|
